
Vol 11 No 1 2026-6

IMPROVEMENTS IN THE SOLUBILIZATION OF THE TT-P0-LS CHIMERIC PROTEIN, A SUBUNIT VACCINE CANDIDATE AGAINST SEA LICE ACTIVE INGREDIENT OF THE SALVAC DEL NORTE VACCINE CANDIDATE
Rutdali María Segura Silva 1, Yunier Luis Paneque Díaz 2 , Mirlleys Peláez Sánchez 3, Nemecio González Fernández4, Alain Moreira Rubio5, Yamila Carpio González6, Amaury Pérez Sánchez 7
1 Centro de Ingeniería Genética y Biotecnologia de Camagüey, Camagüey, Cuba;
ruthdaly.segura@cigb.edu.cu. https://orcid.org/0000-0003-2821-154X
2 Centro de Ingeniería Genética y Biotecnologia de Camagüey, Camagüey, Cuba;
yunier.paneque@cigb.edu.cu. https://orcid.org/0000-0001-6196-0425
3 Centro de Ingeniería Genética y Biotecnologia de Camagüey, Camagüey, Cuba;
mirlleys.pelaez@cigb.edu.cu. https://orcid.org/0009-0003-6190-111X
4 Centro de Ingeniería Genética y Biotecnologia de Camagüey, Camagüey, Cuba;
nemecio.gonzalez@cigb.edu.cu. https://orcid.org/0000-0002-6996-0013
5 Centro de Ingeniería Genética y Biotecnologia de Camagüey, Camagüey, Cuba;
alain.moreira@cigb.edu.cu. https://orcid.org/0000-0003-3293-9410
6 Centro de Ingeniería Genética y Biotecnologia de La Habana, La Habana, Cuba;
yamila.carpio@cigb.edu.cu. https://orcid.org/0000-0002-6238-3663
7 Universidad de Camagüey, Camagüey, Cuba
https://orcid.org/0000-0002-0819-6760
* Correspondence: amaury.perez84@gmail.com
https://doi.org/10.70373/RB/2026.11.01.6
ABSTRACT
Sea louse is one of the marine pathogens that cause the greatest impact on the salmon industry. The chimeric protein TT-P0-Ls, the active ingredient of a vaccine candidate for the control of this entity, is expressed in Escherichia coli, in the form of inclusion bodies. During the protein extraction and refolding process, the stage with the greatest losses is solubilization, which affects its overall efficiency. The objective of the work was to establish the operating conditions in the solubilization stage to increase the yield of the TT-P0-Ls protein. Thus, after establishing the buffer for solubilization, experiments were designed and modeled using the Design Expert program, varying the concentration of dead cell debris, the solubilization time, and the pH of the buffer. Decreasing the concentration of the ruptured cream from 30 to 36 g/L, with a pH of 8 – 9, would achieve an increase in the concentration of the solubilized protein of interest of 2.47 times.
Keywords: sea lice; solubilization; TT-P0-Ls protein; purity; experimental design.
INTRODUCTION
Aquaculture is anticipated to be a key factor in the shift towards healthier and more sustainable diets, necessitating changes toward foods that have lower environmental impacts while meeting dietary requirements 1.
Atlantic salmon (Salmo salar), hereafter just salmon, is a particularly interesting species to study for several reasons. It ranks as one of the top aquaculture species, exhibiting production growth that surpasses that of the overall aquaculture sector. This is to a large extent due to the fact that salmon producers are in the forefront in a number of productivity enhancing categories, such as advancements in production technology and development of the supply chain 2.
Farmed Atlantic salmon constitutes 2% of the worldwide aquaculture sector, with factors influencing productivity growth, such as investments in research, recognized for transforming it into a high-value, global food commodity 1.
From the early 1970s until 2012, the global production of farmed salmon grew at an annual rate of around 23%. The remarkable success of salmon farming is primarily due to advancements in technology, increasing demand, and immediate globalization 3.
Salmon is cultivated in a limited number of countries, yet it faces various economic shocks while being geographically dispersed and subjected to significant differences in biophysical conditions. In 2015, five nations accounted for 95.6% of the production. Norway led the way with a share of 55.3%, followed by Scotland (7.6%) and the Faroe Islands (3.3%) within Europe. The second largest producer, Chile, contributed 25.4% from South America, while Canada added 6% from North America. The remaining 4.4% was distributed among eight countries that face significant constraints on their production capacity due to the availability of suitable sites 2.
Salmon farming faces production risks from biophysical factors such as fluctuations in seawater temperature, harmful algae, sea lice, and various salmon diseases, which in turn influence the volatility patterns of salmon prices 3.
The family Caligidae (Crustacea), which encompasses over 450 species spread across more than 30 genera, includes the salmon louse Lepeophtheirus salmonis, commonly referred to as “salmon lice.” This parasite is frequently found among salmonids and is a part of the marine ecosystem in the Northern Hemisphere 4.
The life cycle of Lepeophtheirus spp. is composed of eight stages: two planktonic nauplii, one infective copepod, two attached chalimus, two mobile pre-adult, and one adult male and female stage. The chalimus stage is particularly crucial for vaccine development, as lice are believed to first engage with their host during this phase through the formation of a frontal filament and subsequent blood feeding, followed by a transition into the adult stage 5.
Sea lice parasitize salmon during the marine stages of their life cycle, in both wild and farmed salmon, by attaching to their skin near the gills and fins 6 and feeding on skin, mucus, and blood, which leads to wounds, stress, and diminished health, potentially causing secondary infections with other pathogens 4.
In the salmon farming sector, the salmon louse has increasingly become a concern due to the abundance of hosts, which promotes ongoing re-infestation and the transfer of lice between farming sites 7.
Lice infestations in aquaculture have a considerable economic and ecological impact on a global scale 8. For instance, in 2018, the expenses related to combating the lice L. salmonis in the production of 1.2 million tons of salmon in Norway were estimated at € 520 million, with a minimum cost of € 0.5 per kilogram of fish 9. Other researchers 8 indicate that in Norwegian aquaculture, the loss of biomass growth due to lice per production cycle ranges from 3.62% to 16.55%, leading to an annual economic loss surpassing US$ 436 million.
Addressing the costly and growing issue of lice requires a diverse array of initiatives and advancements specifically focused on lice control. Innovations can emerge through multiple approaches, which can be categorized as medicinal (such as treatments involving hydrogen peroxide and diflubenzuron), biological (like the use of cleaner fish that consume lice and breeding programs aimed at developing salmon resistant to lice), and mechanical (related to production techniques and equipment). Among these possibilities, mechanical and biological innovations are currently viewed as the most promising, but it is only the mechanical approach that has seen significant targeted government support through policy measures designed to encourage innovation 9.
Farmers utilize two primary strategies to address lice pressure: preventive and reactive treatments. Preventive measures focus on minimizing exposure between infective copepodids and fish, while reactive measures concentrate on eliminating lice that are attached or mobile on the host 8.
Up until 2015, chemical treatment was the preferred approach; however, the emergence of resistance to most chemicals has made this treatment ineffective 4. A significant issue is the resistance developing to the pesticides currently approved, leading to increased usage of these chemicals and the consequent spread of resistance within louse populations, creating a detrimental cycle 7. In recent years, several non-medicinal treatments have emerged as alternatives, but these have resulted in new challenges, including higher post-treatment mortality rates and ethical dilemmas concerning the use of cleaner fish. Consequently, the lice issue has worsened, highlighting the need for innovative control strategies, preferably preventative measures like vaccination 4.
As stated by 10, vaccination is increasingly regarded as an effective and sustainable strategy for preventing a variety of bacterial and viral illnesses. Other authors 6 sustain that vaccination against salmon lice could serve as a significant alternative, as it is well-established that fish vaccines have substantially reduced the reliance on drugs, particularly antibiotics, for treating various fish diseases.
A paper 4 suggests that when searching for vaccine candidates targeting lice, two key criteria must be met for a successful result: (1) the antigens should be essential for the survival, development, or interaction of lice with their host, and (2) they must be presented in a mode that stimulates protective immune responses in the host.
Along with 11, a functional vaccine for sea lice would be highly beneficial and would serve as a preventative measure rather than a treatment option. Nevertheless, creating vaccines for ectoparasites remains a significant technical challenge 12.
In the case of ectoparasites, the significant physical separation between the pathogen and its host further complicates the effectiveness of vaccination. Since the ectoparasite primarily resides externally on the host, it is able to conceal a considerable portion of its constituents (i.e., potential antigens) from the host immune system, effectively putting these antigens “out of reach” of the host’s defense mechanisms 13. If these concealed antigens could be made “visible” to the host immune system, it could potentially trigger a specific immune response that provides protection against them 12.
Numerous studies have been conducted that test or analyze vaccine candidates aimed at controlling sea lice infestations in salmon. In one study 14, a novel gene (denoted as my32) from Caligus rogercresseyi was characterized, which has the highest identity with the Lepeophtheirus salmonis gene akirin-2. To assess the gene’s function, an RNA interference experiment was carried out, resulting in a decrease in the number of ectoparasites on fish within the my32-dsRNA treated group. The recombinant my32 protein was utilized in a vaccination challenge trial to assess its effectiveness in providing protection against sea lice infestations. Notable reductions in the parasite count per fish were recorded at 24 days after the challenge.
In another study 15, the effectiveness of three sea lice vaccines was investigated against the initial stages of infestation, focusing on the transcriptome changes of immunized Atlantic salmon. In this research, three experimental groups of S. salar (Atlantic salmon) received vaccinations with recombinant proteins: Peritrophin (prototype A), Cathepsin (prototype B), and a combination of both (prototype C). The sea lice infestation was assessed during the chalimus I-II stages, which are the early infective phases, 7 days post-infestation. The results indicated that vaccine prototypes A, B, and C led to reductions in parasite burden of 24%, 44%, and 52%, respectively, compared to the control group.
In a recent study 4, the roles of two heme peroxidases from salmon louse (LsPxtl-1 and LsPxtl-2) were evaluated regarding their significance for the parasite’s development and their potential as vaccine candidates. LsPxtl-1 possesses two heme peroxidase domains and is expressed in ovaries and gut, whereas LsPxtl-2 encodes one domain and contains N-terminal signal peptide and an Eph receptor ligand-binding domain. The results of this investigation indicate that LsPxtl-1 is crucial for the development of the parasite; however, the vaccination formulations and methods tested did not markedly decrease lice infestations.
Similarly 6, a vaccine candidate composed of a peptide derived from ribosomal protein P0 was assessed for its effectiveness after L. salmonis infestation in a laboratory environment. The sampling results demonstrated the vaccine candidate’s promising potential when administered intraperitoneally to the host, as it led to a reduction in ectoparasite numbers, lower counts of adult female lice, diminished fecundity, and with greater presumptive effect in F1 lice generation.
Likewise, 11 compared the chimeric P0-my32 protein with the P0 peptide fused to the molecular adjuvant TT regarding immunogenicity and the ability to reduce lice infestation in Atlantic salmon after the experimental challenge. Both P0-my32 and TT-P0 proteins were expressed in E. coli, purified, and formulated with Montanide® ISA50 V2 as an adjuvant. A prime-boost vaccination strategy was implemented, and challenges were conducted at 2, 5, and 11 months following vaccination to examine the onset of immunity and the duration of protection. The findings indicate that vaccination may help diminish lice infestations in Atlantic salmon; however, further studies are necessary to validate effectiveness under field conditions.
The immunogenic properties and protective effects of a vaccine formulation based on a recombinant protein from the salmon lice gut (P33) against L. salmonis infestation in Atlantic salmon was investigated through a laboratory trial 12. The results of this research indicated that the P33 vaccine can offer some degree of protection against both immature and adult salmon lice infestations, with the degree of protection appearing to be dependent on the vaccine dosage, where higher doses led to lower rates of parasitic infestation.
Another study 5 demonstrated the possibility of developing a cost-effective synthetic peptide vaccine based on the secretions injected into the host by the parasite. The protein peroxiredoxin-2, obtained from the blood of Atlantic salmon affected by lice, was believed to contribute to antioxidative processes that aid the parasite’s feeding. In this study, vaccinating Atlantic salmon with an experimental vaccine (peptide purity of 70%) provided superior protection against C. rogercresseyi (92%) compared to L. salmonis (60-70%).
Other examined approaches for controlling sea lice include the application of fine calcium oxide (CaO) particles on the planktonic stages of salmon lice under controlled lab conditions. The findings of this research showed that exposure to CaO (for 10 minutes at 0.2 g/L) decreased the number of nauplii progressing to the copepodid stage by 60-70% by reducing both survival and molting, while treatment with CaO (for 10 minutes at 0.6 g/L) resulted in up to 90% mortality of free-living copepodids 8.
In the study by 16, an identified non-host semiochemical, 2-aminoacetophenone (2-AA), isolated from turbot (Scophthalmus maximus) was used in the field to reduce sea lice prevalence and infective capability. Field experiments took place at a research fish farm located in north-west Scotland to assess the impact of 2-AA on the population of L. salmonis on Atlantic salmon (S. salar) hosts.
The creation of a vaccine candidate that targets conserved proteins shared between the pathogen and the host has a significant drawback: the high similarity can lead to the development of tolerance or the production of autoantibodies in the host. To address this issue, peptide-based vaccines have been formulated that demonstrate notable efficacy 17; for example, a vaccine developed targeting ticks that features a 20-amino acid peptide derived from the acidic ribosomal protein P0 of Rhipicephalus sp. ticks 18,19.
With respect to the implementation and evaluation of downstream processes conducted in Cuban research facilities for antigen purification aimed at developing vaccine candidates against sea lice, two studies are reported.
In the first study 20, a compact purification method was developed to isolate the MY32/Ls protein, analyzing various alternatives to define the steps for rupture, chromatography, and renaturation, in order to obtain the production of the active pharmaceutical ingredient (API). Due to techno-economic advantages, a chemical rupture process was established using 8 mol/L urea for 1 hour, which was 20 % cheaper than mechanical rupture with glass beads. For washing and elution of the protein in the chromatographic step, pH and imidazole were evaluated. The specifications for the API produced in this study met the necessary standards for formulating the new vaccine against sea lice.
In the second study 21, the purification procedures for the MY32/Ls protein were designed and optimized to obtain the API against sea lice. This investigation utilized non-chromatographic purification methodologies based from existing literature to establish conditions for rupture, washing, solubilization, and refolding. The API yielded in this research demonstrated a superior immune response (IgG) compared to the positive control and exhibited higher purity.
In 17 the chimeric protein TT-P0-Ls (TT- The promiscuous T cell epitopes of tetanus toxin. P0- A 35 aminoacids peptide between aminoacids 267 and 301 of the sequence of P0 of L. salmonis. Ls- L. salmonis) was obtained, with high immunogenicity, by recombinant DNA methods, and expressed in the form of inclusion bodies in the E. coli strain (BL21, DE3). The objective of the present work is to increase the purity and recovery of the TT-P0-Ls protein in the solubilization stage.
MATERIALS AND METHODS
- coli propagation stage
A plate was streaked containing solid LB medium with the E. coli strain BL21 (DE3) transformed with the PET28aTT-P0-Ls plasmid 17. The streaked plate was incubated for 24 h at 37 °C in a thermostated shaker (RETOMED, Cuba). From the grown colonies, a sample was taken to inoculate a 2 L Erlenmeyer flask containing 500 mL of non-inducible culture medium. Also, 15 mL of 40% glucose and 500 µL of kanamycin was added to each Erlenmeyer at a final concentration of 50 µg/mL and then incubated in a thermostated shaker (RETOMED) for 16 h at 37 °C with constant stirring at 250 rpm.
Fermentation Stage
Fermentations were carried out in a fermentor (Marubishi) with 50 L of autoinduction medium with lactose, to which 3 L of the previously prepared inoculum was added. Fermentations last for 24 h. A temperature of 37 °C is maintained throughout the fermentative process while the pH is controlled in 7.0 by adding 25% ammonia or 20% orthophosphoric acid. During the fermentation process, the foam was controlled with Glanapon DG-158 antifoam prepared at 20%. Constant agitation was maintained in 500 rpm and aeration in 75 L/min.
Harvest Stage
The culture was centrifuged for 30 min at 3,000 rpm using batch centrifuges (CEPA) to eliminate the remains of culture medium and ensure that in the subsequent stages there are no traces of the antibiotic used during the preceding stages. The culture was kept at a temperature of – 20 °C in a refrigerated room.
Recovery Stage
The washed and concentrated cream was thawed and homogenized for 20 min using a dispersing agitator (Polytron) in a disruption buffer (300mM NaCl, 100mM sodium phosphate buffer, pH 6). The rupture was performed with the ball mill (DynoMill) carrying out a total of 5 passes. After the rupture operation, the washing solution (300 mM NaCl, 10 mM Tris, pH 6) and the ruptured biomass are added. The resulting mixture is homogenized for 10 min using a dispersing agitator (Polytron) at 10,000 rpm. After homogenization, the solid and liquid phases are separated in a batch centrifuge (CEPA), where the sample is centrifuged at 12,000 rpm for 1 h at 4 °C.
Experimental design for TT-P0Ls solubilization
A factorial design was developed using the Design Expert 8.0.6 program (Stat-Ease, Inc.). Three independent variables that influence protein solubility were taken into account: time (1, 6.5, 12 and 15 h); pH of the medium (pH of the solubilization solution) (6; 7; 8.5; 10 and 11) and concentration of the ruptured cells (1; 19.77; 30; 45; 60 and 70.23 g/L), resulting in 15 experimental runs (Table 1).
Table 1. Factorial design of the experiments obtained using the Design Expert software.
| Run | Factor A:
Solubilization time (h) |
Factor B:
pH of the solubilization solution |
Factor C:
Concentration of ruptured cells (g/L) |
| 1 | 6.50 | 8.50 | 70.23 |
| 2 | 6.50 | 8.50 | 19.77 |
| 3 | 1.00 | 7.00 | 60.00 |
| 4 | 12.00 | 10.00 | 60.00 |
| 5 | 1.00 | 10.00 | 30.00 |
| 6 | 6.50 | 11.00 | 45.00 |
| 7 | 15.75 | 8.50 | 45.00 |
| 8 | 12.00 | 10.00 | 30.00 |
| 9 | 6.50 | 8.50 | 45.00 |
| 10 | 0.00 | 8.50 | 45.00 |
| 11 | 6.50 | 5.98 | 45.00 |
| 12 | 12.00 | 7.00 | 60.00 |
| 13 | 1.00 | 10.00 | 1.00 |
| 14 | 12.00 | 7.00 | 30.00 |
| 15 | 1.00 | 7.00 | 30.00 |
Variants of the solubilization solution (10 mM Tris, 10 mM Imidazole, 8 M Urea) were prepared at different pH values according to the experimental design described in Table 1, using hydrochloric acid 1M to decrease the pH, and Tris 1M to increase the pH, and employing a dispersing homogenizer (IKA T25). Then, the solution was mixed with the washed cream containing the different concentrations of broken cells (Table 1). The mixtures were homogenized with the help of a magnetic stirrer (Heidolph MR) and incubated at 37 °C in a thermostated shaker (RETOMED) operating at 150 rpm for 15 h. Samples were taken at different solubilization times according to the experimental design (Table 1). After the solubilization time had elapsed, the samples were centrifuged at 10,000 rpm for 30 min at 4 °C.
Determination of total protein concentration and percentage of purity of the TT-P0-Ls protein
Once the centrifugation was completed, 1 mL of the extraction supernatant was taken and the total protein concentration was determined by the bicinchoninic acid method 22. The clarified protein was also analyzed by the SDS-PAGE electrophoresis method 23 in a 15% acrylamide/bisacrylamide gel, with the aim of observing the purity and the presence of the band at the height of 7 – 15 kDa. The remaining supernatant containing the solubilized protein was aliquoted and stored at 4 °C until use.
From the values of the response variables considered in this study (Concentration of total proteins), an ANOVA analysis was carried out using the Design Expert program to know the independent variables that influence the most in the solubilization of the TT-P0-Ls protein.
RESULTS
Figure 1 shows the numerical values of the response variable considered in this study, that is, concentration of total proteins (in mg of total proteins/g of biomass) for each of the experimental runs included in the design of experiments.
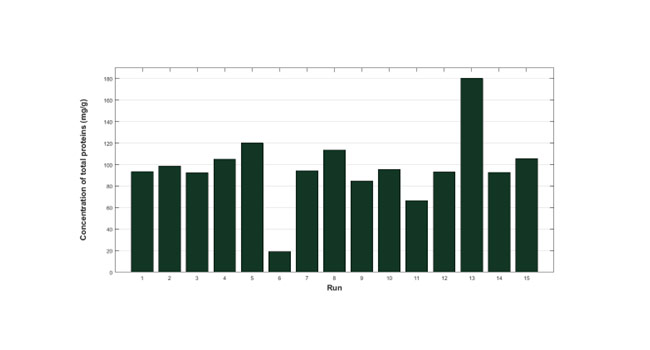 |
| Figure 1. Results of total protein concentration according to the experimental design. |
Table 2 presents the results of the ANOVA analysis performed by the Design Expert program.
Table 2. ANOVA analysis performed in the Design Expert program.
| Source | Sum of squares | Degree of freedom | Mean of variables | F Value | P Value
(Prob > F) |
| Model | 12,497.94 | 9 | 1,388.66 | 2.72 | 0.14 |
| A-Time | 2.10 | 1 | 2.10 | 4.11E-003 | 0.95 |
| B-pH | 288.80 | 1 | 288.80 | 0.56 | 0.48 |
| C-Conc. | 459.57 | 1 | 459.57 | 0.90 | 0.38 |
| AB | 279.76 | 1 | 279.76 | 0.55 | 0.49 |
| AC | 355.99 | 1 | 355.99 | 0.70 | 0.44 |
| BC | 258.15 | 1 | 258.15 | 0.51 | 0.50 |
| A2 | 429.27 | 1 | 429.27 | 0.84 | 0.40 |
| B2 | 1,129.16 | 1 | 1,129.16 | 2.21 | 0.19 |
| C2 | 598.09 | 1 | 598.09 | 1.17 | 0.32 |
| Residue | 2,555.82 | 5 | 511.16 | ||
| Cor Total | 15,053.76 | 14 |
Figure 2 graphically shows the result of the ANOVA analysis.
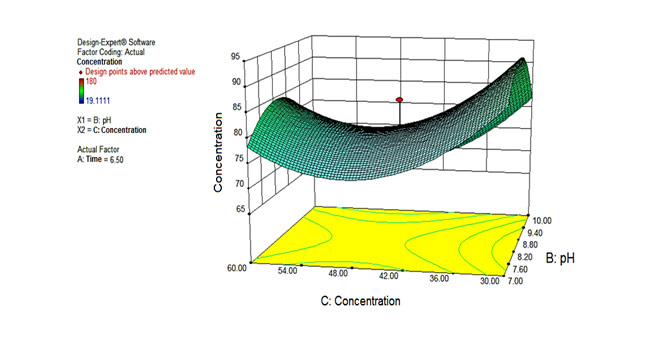 |
| Figure 2. Result of ANOVA analysis in 3D coordinates. X axis: concentration of dead cells in solubilization. Y axis: pH of solubilization. Z axis: Protein concentration in the solubilization supernatant. The time to elaborate the graph was set at 6 h and 30 min (6.50 h). |
Figure 3 presents in the image on the left the SDS-Page 15% of the proteins used according to the design of the experiment, containing in the first lane the positive control of the protein TT-P0-Ls (C+), and in the lanes of the 1 to 10 the different experimental runs. Likewise, the image on the right shows the results of the purity (%) of the solubilized protein for each of the experiments carried out.
 Figure 3. Graphics related to: Figure 3. Graphics related to:
a) SDS-PAGE 15% under reducing conditions. b) Percent purity of the TT-P0-Ls protein in the solubilization supernatant at the different experimental points. |
|
Table 3 exhibits the results of the concentration of total proteins in the solubilization stage in three different batches.
Table 3. Results of the concentration of total proteins in the solubilization stage in three different batches.
| Batch code | Concentration
(mg total proteins/g cell debris) |
| 68N.2107 | 44.33 |
| 68N.2108 | 12.89 |
| 68N.2109 | 20.36 |
Figure 4 displays the SDS-PAGE 15% under reducing conditions of the samples generated in the rupture stage.
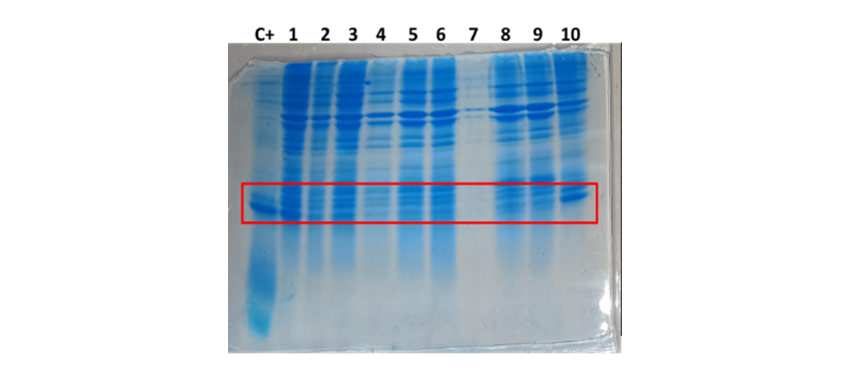 |
|
| Figure 4. SDS-PAGE 15% under reducing conditions of the samples generated in the rupture stage. From left to right 1: Molecular weight standard (PPM), 2: TT-P0-Ls protein positive control (C+), 3: Sample from the last rupture pass (R1), 4: Wash supernatant (R2), 5: Remains of dead cells (R3), 6: TT-P0-Ls protein after solubilization process (R4). | |
Figure 5 shows the graphics related to SDS-PAGE 15% under reducing conditions and the percent purity of the TT-P0-Ls protein in the cellular debris.
 Figure 5. Graphics related to: Figure 5. Graphics related to:
a) SDS-PAGE 15% under reducing conditions. b) Percent purity of the TT-P0-Ls protein in the cellular debris. |
|
DISCUSSION
The inclusion bodies produced in E. coli consist of a dense packing of denatured protein molecules in the form of particles. The renaturalization process, after they are extracted from the inclusion bodies, is arduous and usually has poor recovery and increases the production costs 24.
The use of factorial design assisted by computer programs allows optimizing the number of experiments to be carried out, by proposing the best test variants considering the variables that most affect the processes 25.
According to the results of Table 2 (ANOVA analysis), time is the parameter that least influences in the increment in the concentration of solubilized protein due to its high value (p = 0.95), therefore it is concluded that time does not affect the solubilization process of total proteins.
In this study, the concentration of ruptured cells is the variable that most influences the concentration of solubilized proteins, although there are no significant differences with the other variables (p=0.38)
It can be observed from Figure 2 that the pH and the concentration of dead cell residues are the parameters that most influence the concentration of solubilized proteins. This figure was elaborated for a time of 6 h and 30 min (6.50 h). Figure 2 also exposes that the optimal conditions of the pH parameter are in the range of 8-9, while the best results of the parameter concentration of broken cells are in the range of 30-36 mg total proteins/g biomass. This is because the TT-P0Ls protein is more stable at pH 8 to 9 and does not precipitate when separating the dead cells from the protein suspension.
A study 24 reports that pure recombinant human growth hormone (r-hGH) inclusion bodies were solubilized at different pHs in 100 mM Tris buffer (pH 3–13) and percent solubilization of r-hGH was monitored. The paper also indicated that the solubilization of r-hGH from inclusion bodies was observed by increasing the pH from 6 to 12.5. Higher solubilization of r-hGH from inclusion bodies was observed by incorporating 2M urea in 100 mM Tris buffer at pH 12.5. Further addition of urea in 100 mM Tris buffer at pH 12.5 did not further increase solubilization of r-hGH from the inclusion bodies. In 100 mM Tris buffer at pH 12.5 containing 2M urea, a maximum of 6 mg/ml of r-hGH were solubilized from the inclusion bodies. Use of 2M urea did not unfold the protein completely and preserved the native-like secondary structure. Finally, these authors stated that a combination of alkaline pH and 2M urea destabilized both the ionic and hydrophobic interactions which are the major cause of protein aggregation in inclusion bodies of human growth hormone.
Also, another study 26 indicates that the preparation of good conditions for forming secondary structure of inclusion bodies during the solubilization step is momentous to decrease aggregation. Ionic and hydrophobic interactions are the 2 major factors to induce aggregation, while the presence of urea at the low concentration or the change of pH, particularly alkaline pH, in the solubilization step help to restore the secondary structures of inclusion bodies following with a better refolding of protein along with high levels of bioactivity. Therefore, using the mild process to solubilize the inclusion body and also select the protocol to dilute the concentration of proteins and remove impurity could be helpful to decrease the aggregation and enhance the yield of bioactive recombinant proteins.
Likewise, a research was carried out 27 to investigate whether altering the pH might improve inclusion body resolution at lower urea concentrations as milder concentrations of denaturants ensure preservation of the proteins’ secondary structure during the solubilization process. The results indicated that increasing the pH value of the 2 M urea buffer from 8.5 to 10.5 significantly improved the solubilization, indicating that solubility of TrxmGH inclusion bodies was pH-dependent with optimal solubilization observed at a pH value of 10.5. This study also specifies that developing an efficient method to extract biologically active proteins from inclusion bodies is often inevitable for certain proteins. For effective inclusion body solubilization and recovery, optimization of multiple parameters is required, such as the denaturant concentration, pH values, and the refolding method. However, ‘mild’ solubilization conditions, such as lower denaturant concentrations and alkaline pH, ensure better recovery of the proteins in its conserved native structure, but also help to avoid protein aggregations during the refolding process.
Singh et al. 28 denotes that non-classical inclusion bodies (inclusion body aggregates which are biologically active) have been reported to be very sensitive to pH at which they are purified, as use of high pH can lead to solubilization of protein molecules during purification. It also declared that mild solubilization processes using alkaline pH, high pressure, detergents, organic solvents and low concentration of chaotropes have been used for recovery of bioactive proteins from inclusion bodies. In most of the cases, improvements in inclusion body isolation and use of modern refolding methods in combination with mild solubilization improved the overall recovery of bioactive proteins. Likewise, the use of buffers with extreme pH has also been reported as a mild solubilization method. High pH (>12) buffer in combination with 2 M urea has been used successfully for solubilization of inclusion bodies. High pH buffer has been described to improve refolding yield by retaining native-like secondary structures in solubilized state.
Finally, a study 29 indicates that it has been widely reported that growth hormone inclusion bodies of different species expressed in E. coli can be solubilized by alkaline pH. These authors report that the solubilization of the recombinant human growth hormone r-hGH from inclusion bodies was observed by increasing the pH from 8 to 12.5. High alkaline pH (> 12.5), even though it helped in solubilizing r-hGH from inclusion bodies, resulted in extensive degradation of r-hGH. A maximum of 2 mg/ml of r-hGH was solubilized in 100 mM Tris buffer at pH 12.5 without the addition of urea or guanidine hydrochloride. These authors also declare that charge distribution provided by high alkaline pH, along the protein chain, was responsible for higher solubilization of r-hGH from inclusion bodies. This suggested that pH has a crucial role in destabilizing the inclusion body aggregation. Changing the charge distribution along the protein molecule by changing the pH generally affects the protein stability and may lead to an unfolding of the native protein. Finally, they indicate that the use of 2 M urea at alkaline pH improved r-hGH solubilization from inclusion bodies without disturbing the existing native-like secondary structure of the proteins.
The concentration of dead cells in the cell suspension is the parameter that has the most influence because the denaturing agents achieve total or partial rupture of the inclusion bodies. Solubilization must be able to break the protein packaging of the inclusion bodies and solubilize the proteins to their minimum structural state or a state that allows their subsequent purification. Most reports in the literature used high concentrations of the denaturing agent (between 6 and 8 mol/L) and protein concentrations in the solubilization step between 1 and 10 mg/mL 30-36.
Singh et al. 28 state that the process of protein recovery from inclusion bodies involves four steps:
- Purification of inclusion bodies to homogeneity (careful lysis of cells, purify inclusion bodies by detergent washing/ultracentrifugation),
- Solubilization of inclusion bodies using mild solubilization agent (alkaline pH/hydrostatic pressure/organic solvent based buffers/2-3 M urea),
- Refolding of the solubilized proteins (refolding at high protein concentration by pulsatile renaturation/on column refolding/urease mediated refolding) with optimal refolding buffer, and
- Purification of the refolded protein. These steps can be optimized to recover high amount of bioactive protein from the inclusion bodies of coli.
It’s worth mentioning that in the case of our research, detergents were not used in the solubilization step due to regulations issued by the International Cooperation on Harmonisation of Technical Requirements for the Registration of Veterinary Medicinal Products (VICH).
In Figure 3a the presence of the TT-P0-Ls protein can be observed in the supernatant obtained in the solubilization stage (indicated in the box with red borders), while in Figure 3b it is observed that the greatest protein purity is achieved at pH 8.5, with a decrease in purity occurring at a pH greater than 10.
In the three batches previously produced the concentration of total proteins in the solubilization stage did not have a stable behavior (Table 3), obtaining a low purity (14%) and the protein TT-P0-Ls was present in the cellular debris resulting from solubilization (Figure 4).
Figure 5 shows that there is a decrease in the TT-P0-Ls protein band in the cellular debris starting at a pH value of 8.5. Also, the purity of the TT-P0-Ls protein band in the cellular debris (and therefore the amount of this protein) increases at the extreme values of pH (6-7 and 11), while decreases at pH 8.5.
The subsequent execution of three experimental batches allowed more than 38 mg of total proteins per g of cellular debris to be obtained, with 31% purity at 25 ± 3 ºC.
With the new process established in this work, the solubilization time would be reduced from 3 h to 1 h, obtaining an average concentration of total proteins of 95.55 mg/g of cellular debris and a purity of 40-50% at a pH of 8.5-9.
CONCLUSIONS
In the current study, up to 50% purity of the protein of interest in the supernatant and an increase in protein of interest of 2.47 times was achieved using a concentration of ruptured biomass in the range 30 – 36 g/L, at pH between 8 and 9, for 1 h at room temperature, while the amount of the protein of interest in the precipitate is reduced, after the solubilization was carried out. These results set a good start point for the development of an efficient biotechnological process to obtain TT-P0-Ls antigen for further use as therapeutic vaccine against sea lice.
Author Contributions: Conceptualization, R.M,S.S., Y.L.P.D. and Y.C.G.; methodology, R.M.S.S., Y.L.P.D. and M.P.S.; software, Y.L.P.D. and M.P.S.; validation, N.G.F., A.M.R. and Y.C.G.; formal analysis, R.M.S.S. and A.P.S; investigation, R.M.S.S., Y.L.P.D., M.P.S., and Y.C.G.; resources, A.M.R. and A.P.S; data curation, Y.L.P.D., Y.C.G. and N.G.F.; writing—original draft preparation, Y.L.P.D., M.P.S. and A.P.S.; writing—review and editing, R.M.S.S., Y.C.G. and N.G.F.; visualization, A.M.R. and A.P.S..; supervision, R.M.S.S., Y.L.P.D. and Y.C.G..; project administration, R.M.S.S. and Y.C.G.; funding acquisition, R.M.S.S. and Y.L.P.D. All authors have read and agreed to the published version of the manuscript.
Funding: This research received no external funding.
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Acknowledgments: The authors wish to thank to the scientists and technicians of the Animal Biotechnology Department of the Centre for Genetic Engineeering and Biotechnology of Havana for their support and collaboration.
Conflicts of Interest: The authors declare no conflict of interest.
REFERENCES
- Robinson, J.P.W.; Fernandes, J. The hidden role of market-making in the rise of farmed salmon. Environmental Research Letters, 2025, 20, 084061. https://doi.org/https://doi.org/10.1088/1748-9326/adec05
- Iversen, A.; Asche, F.; Hermansen, Ø.; Nystøyl, R. Production cost and competitiveness in major salmon farming countries 2003–2018. 2020, 522, 735089. https://doi.org/https://doi.org/10.1016/j.aquaculture.2020.735089
- Zhang, D.; Fang, Y.; Liu, Y. Import demand under price and exchange-rate uncertainties: The case of U. S. Atlantic salmon imports. Aquaculture. 2023, 574,739712. https://doi.org/https://doi.org/10.1016/j.aquaculture.2023.739712
- Gislefoss, E.; Gamil, A.A.A.; Øvergard, A-C.; Evensen, Ø. Identification and characterization of two salmon louse heme peroxidases and their potential as vaccine antigens. iScience. 2023, 26, 107991. https://doi.org/https://doi.org/10.1016/j.isci.2023.107991
- Johny, A.; Ilardi, P.; Olsen, R.E.; Egelandsdal, B.; Slinde, E. A Proof-of-Concept Study to Develop a Peptide-Based Vaccine against Salmon Lice Infestation in Atlantic Salmon (Salmo salar). Vaccines. 2024, 12, 456. https://doi.org/https://doi.org/10.3390/vaccines12050456
- Swain, J.K.; Carpio, Y.; Johansen, L-H.; Velazquez, J.; Hernandez, L.; Leal, Y. Impact of a candidate vaccine on the dynamics of salmon lice (Lepeophtheirus salmonis) infestation and immune response in Atlantic salmon (Salmo salar). PLoS ONE. 2020, 15, e0239827. https://doi.org/https://doi.org/10.1371/journal.pone.023982
- Sandlund, L.; Nilsen, F.; Male, R.; Grotmol, S.; Kongshaug, H.; Dalvin, S. Molecular characterisation of the salmon louse, Lepeophtheirus salmonis salmonis (Krøyer, 1837), ecdysone receptor with emphasis on functional studies of female reproduction. International Journal for Parasitology. 2015, 45, 175-85. https://doi.org/http://dx.doi.org/10.1016/j.ijpara.2014.10.003
- Ciani, E.; Stormoen, M.; Antonsen, S.I.; Nilsen, F.; Jørgensen, E.H.; Komisarczuk, A.Z.; Optimization of calcium oxide treatment against salmon louse (Lepeophtheirus salmonis). A controlled laboratory study. Aquaculture Reports. 2025, 43, 102894. https://doi.org/https://doi.org/10.1016/j.aqrep.2025.102894
- Rosendal, G.K.; Olesen, I. Overcoming barriers to breeding for increased lice resistance in farmed Atlantic salmon: A case study from Norway. Aquaculture. 2022, 548, 737574. https://doi.org/https://doi.org/10.1016/j.aquaculture.2021.737574
- Mechlaoui, M.; Nordstrand, E.; Strandskog, G.; Jensen, I.; Seternes, T. Vaccinated Atlantic salmon (Salmo salar) maintain a specific antibody response throughout the seasonal fluctuations of a full commercial production cycle in sea: a case study. Aquaculture. 2025, 595, 741536. https://doi.org/https://doi.org/10.1016/j.aquaculture.2024.741536
- Rodríguez, A.; Gadan, K.; Pérez, L.; Evensen, Ø.; Estrada, M.P.; Carpio, Y. Prime-boost vaccination with chimeric antigens adjuvanted in Montanide™ ISA50 V2 confers protection against experimental Lepeophtheirus salmonis infestation in Atlantic salmon (Salmo salar). Frontiers in Immunology. 2025, 16, 1570948. https://doi.org/10.3389/fimmu.2025.1570948
- Tartor, H.; Karlsen, M.; Skern-Mauritzen, R.; Monjane, A.L.; Press, C.M.; Wiik-Nielsen, C.; et al. Protective Immunization of Atlantic Salmon (Salmo salar) against Salmon Lice (Lepeophtheirus salmonis) Infestation. Vaccines. 2022, 10, 16. https://doi.org/https://doi.org/10.3390/vaccines10010016
- Allen, J.R. Host resistance to ectoparasites. Revue Scientifique et Technique. 1994, 13, 1287-1303. https://doi.org/http://dx.doi.org/10.20506/rst.13.4.824
- Carpio, Y.; Basabe, L.; Acosta, J.; Rodríguez, A.; Mendoza, A.; Lisperger, A.; et al. Novel gene isolated from Caligus rogercresseyi: A promising target for vaccine development against sea lice. Vaccine. 2011, 29, 2810-2820. https://doi.org/10.1016/j.vaccine.2011.01.109
- Casuso, A.; Valenzuela-Muñoz, V.; Benavente, B.P.; Valenzuela-Miranda, D.; Gallardo-Escárate, C. Exploring Sea Lice Vaccines against Early Stages of Infestation in Atlantic Salmon (Salmo salar). Vaccines, 2022, 10, 1063. https://doi.org/10.3390/vaccines10071063
- Hastie, C.; Wallace, C.; Birkett, M.A.; Douglas, A.; Jones, O.; Mordue, A.J.; et al. Prevalence and infection intensity of sea lice (Lepeophtheirus salmonis) on Atlantic salmon (Salmo salar) host is reduced by the non-host compound 2-aminoacetophenone. 2013, 410-411, 179-183. http://dx.doi.org/10.1016/j.aquaculture.2013.06.035
- Leal, Y.; Velazquez, J.; Hernandez, L.; Swain, J.K.; Rodríguez, A.R.; Martínez, R.; et al. Promiscuous T cell epitopes boosts specific IgM immune response against a P0 peptide antigen from sea lice in different teleost species. Fish and Shellfish Immunology. 2019, 92, 322-330. https://doi.org/https://doi.org/10.1016/j.fsi.2019.06.018
- Rodríguez-Mallon, A.; Encinosa, P.E.; Méndez-Pérez, L.; Bello, Y.; Fernández, R.R.; Garay, H.; et al. High efficacy of a 20 amino acid peptide of the acidic ribosomal protein P0 against the cattle tick, Rhipicephalus microplus. Ticks and Tick-borne Diseases. 2015, 6, 530-537. http://dx.doi.org/10.1016/j.ttbdis.2015.04.007
- Rodríguez-Mallon, A.; Fernández, E.; Encinosa, P.E.; Bello, Y.; Méndez-Pérez, L.; Ruiz, L.C.; et al. A novel tick antigen shows high vaccine efficacy against the dog tick, Rhipicephalus sanguineus. Vaccines. 2012, 30, 1782-1789. https://doi.org/10.1016/j.vaccine.2012.01.011
- Perez, C.E.; González, N.; León, L.; Salazar, E.; Pimentel, R.; Pimentel, E.; et al. Design of the purification process by metal-chelate affinity chromatography of a new vaccine antigen for the control of sea lice. Biotecnología Aplicada. 2015, 32, 1211-1216.
- Perez, C.E.; González, N.; Salazar, E.; Pimentel, E.; Carpio, Y.; Limonta, M.; Design and Optimization of a Purification Process for MY32/Ls Protein Solubilizing Inclusion Bodies for a New Vaccine Against Sea Lice. BioProcessing Journal. 2015, 14, 3-13. https://doi.org/10.12665/J141.Heredia
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Frovenzano, M.D.; et al. Measurement of Protein Using Bicinchoninic Acid. Analytical Biochemistry. 1985, 150, 76-85.
- Hames, B.D.; Rickwood, D. Gel Electrophoresis of Proteins: A Practical Approach: IRL Press; 1981.
- Singh, S.M.; Panda, A.K. Solubilization and Refolding of Bacterial Inclusion Body Proteins. Journal of Bioscience and Bioengineering. 2005, 99, 303-310. https://doi.org/10.1263/jbb.99.303
- Collins, L.M.; Dziak, J.J.; Kugler, K.C.; Trail, J.B. Factorial Experiments – Efficient Tools for Evaluation of Intervention Components. American Journal of Preventive Medicine. 2014, 47, 498-504.
- Mirhosseini, S.A.; Latifi, A.M.; Hosseini, H.M.; Seidmoradi, R.; Aghamollaei, H.; Farnoosh, G. The Efficient Solubilization and Refolding of Recombinant Organophosphorus Hydrolyses Inclusion Bodies Produced in Escherichia coli. Journal of Applied Biotechnology Reports. 2019, 6, 20-25. https://doi.org/10.29252/JABR.06.01.04
- Kim, M.; Langley, R.J.; Perry, J.K.; Wang, Y. Recovery of mouse growth hormone from coli inclusion bodies using a mild solubilisation and repeated freeze–thaw approach. Molecular Biology Reports. 2025, 52, 627. https://doi.org/10.1007/s11033-025-10685-y
- Singh, A.; Upadhyay, V.; Upadhyay, A.K.; Singh, S.M.; Panda, A.K. Protein recovery from inclusion bodies of Escherichia coli using mild solubilization process. Microbial Cell Factories. 2015, 14, 41. https://doi.org/10.1186/s12934-015-0222-8
- Patra, A.K.; Mukhopadhyay, R.; Mukhija, R.; Krishnan, A.; Garg, L.C.; Panda, A.K. Optimization of Inclusion Body Solubilization and Renaturation of Recombinant Human Growth Hormone from Escherichia coli. Protein Expression and Purification. 2000, 18, 182-192. https://doi.org/10.1006/prep.1999.1179
- Batas, B.; Chaudhuri, J.B.; Considerations of sample application and elution during size-exclusion chromatography-based protein refolding. Journal of Chromatography A. 1999, 864, 229-236. https://doi.org/10.1016/S0021-9673(99)01030-4
- Clark, E.D.B.; Schwarz, E.; Rudolph, R. Inhibition of Aggregation Side Reactions during in Vitro Protein Folding. Methods in Enzymology. 1999, 309, 217-236. https://doi.org/10.1016/S0076-6879(99)09017-5
- Fahey, E.M.; Chaudhuri, J.B.; Binding, P. Refolding and purification of a urokinase plasminogen activator fragment by chromatography. Journal of Chromatography B. 2000, 737, 225-35. https://doi.org/10.1016/S0378-4347(99)00360-6
- Huxtable, S.; Zhou, H.; Wong, S.; Li, N. Renaturation of 1-Aminocyclopropane-1-carboxylate Synthase Expressed in Escherichia coli in the Form of Inclusion Bodies into a Dimeric and Catalytically Active Enzyme. Protein Expression and Purification. 1998, 12, 305-314. https://doi.org/10.1006/prep.1997.0847
- Misawa, S.; Kumagai, I. Refolding of Therapeutic Proteins Produced in Escherichia coli as Inclusion Bodies. Biopolymers (Peptide Science). 1999, 51, 297-307. https://doi.org/10.1002/(SICI)1097-0282(1999)51:4<297::AID-BIP5>3.0.CO;2-I
- Müller, C.; Rinas, U. Renaturation of heterodimeric platelet-derived growth factor from inclusion bodies of recombinant Escherichia coli using size-exclusion chromatography. Journal of Chromatography A. 1999, 855, 203-13. https://doi.org/10.1016/S0021-9673(99)00660-3
- Tran-Moseman, A.; Schauer, N.; Clark, E.D.B. Renaturation of Escherichia coli-Derived Recombinant Human Macrophage Colony-Stimulating Factor. Protein Expression and Purification. 1999, 16, 181-189. https://doi.org/10.1006/prep.1999.1074
Received: 12 diciembre 2025 / Accepted: 22 febrero 2026 / Published: 15 marzo 2026
Citation: Segura, R; Paneque, Y; Peláez, M; González, N; Moreira, A; Carpio, Y; Pérez A. Improvements in the solubilization of the tt-p0-ls chimeric protein, a subunit vaccine candidate against Sea Lice active ingredient of the Salvac del Norte vaccine candidate. Bionatura. 2026. Volumen 11, No 1. https://doi.org/10.70373/RB/2026.11.01.6
Publisher’s Note: Bionatura stays neutral concerning jurisdictional claims in published maps and institutional affiliations.
Copyright: © 2026 by the authors. Submitted for possible open-access publication under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Vol11 No1 2026

INDEXADA EN
INDEXADA EN
